




Purolite and Repligen have commercially launched a new CH1 affinity resin for the purification of specialized mAbs.
Purolite and Repligen have commercially launched a new CH1 affinity resin for the purification of specialized mAbs.
Product and process understanding are essential to effective resin selection.
The 3M Harvest RC Chromatographic Clarifier, BT500 is the latest addition to the company’s chromatographic clarifier portfolio.

Innovations in downstream processing accelerate development, enhance efficiency, and promote sustainability.

CPC presents a green alternative to chromatography that supports sustainable operations.

The breakthrough approach is commonly used, but the devil is in the details.

Advanced LC technologies can streamline the review process for LC equipment maintenance.

Affinity chromatography resins must perform well under mild elution conditions yet withstand robust cleaning and sanitization protocols.

A thorough knowledge of both the upstream and downstream processes is required to ensure effective removal of residual impurities in biopharmaceuticals.

Enriching process understanding enables robust process controls.
Thanks to technological advances, process chromatography challenges are surmountable, but increasingly complex therapies in the pipeline will demand more efficient processes in the future.

Emerging drug classes are giving rise to challenges in downstream processing, pushing the need for strategizing efficiencies.
Enhancement of resin technology can improve the ability of chromatography to purify the growing landscape of increasingly complex biomolecules.
Thermo Fisher Scientific showcases its new mass spectrometry and chromatography solutions at the ASMS Conference on Oct. 31–Nov. 4, 2021.

Process chromatography in downstream processing has seen steady improvements over the years, but there is still room for improvement.

Mechanistic models provide process understanding for developing robust manufacturing processes and for scale up and tech transfer.

Recent innovations in chromatography resins offer promising advantage in downstream bioprocessing.
Sartorius has expanded in the United Kingdom with the relocation to a new facility in Havant, Hampshire, for downstream processing equipment.
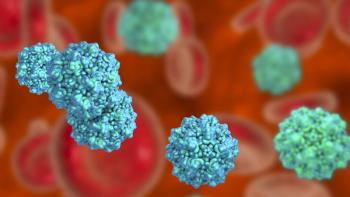
A series of minute virus of mice spiking studies were performed with a monoclonal antibody product and small-scale AEX membrane adsorbers to evaluate a range of critical processing conditions.
The launch of HiScreen Fibro PrismA, the company’s newest product for early mAb purification process development strengthens Cytiva’s new fiber-based Protein A platform.
The transaction will include operations located at different Novasep sites, including its Pompey, France site and sites in the United States, China, and India.
The development of an innovative purification process simplifies downstream processing for biologics.

Establishing an automated inline dilution system can potentially ease bottlenecking delays resulting from higher upstream yields.

Continuous SEC was shown to increase productivity with the same product quality and yield.

The latest advances in downstream technologies include sterile disconnects for single-use manufacturing systems, ligand technologies for purifying RNA and for COVID-19 vaccines, and an automated perfusion system.